低分子量G蛋白質Rap1とエフェクター分子RAPLによるインテグリン接着制御:
インテグリンは細胞の接着を担う細胞表面分子で、接着を迅速に変化させることによって細胞接着だけでなく細胞移動も調節しています。このような他の接着分子にみられないユニークな調節機構は個体発生や臓器形成に重要であるばかりでなく、免疫系における活発なリンパ球や樹状細胞の生体内動態に重要な役割を果たし、また癌の転移機構などにも関与しています。当研究室ではインテグリン接着制御の分子機構を解明し、その生理的役割と破綻による異常を細胞レベルから個体レベルで明らかにすることを主な研究目標としています。
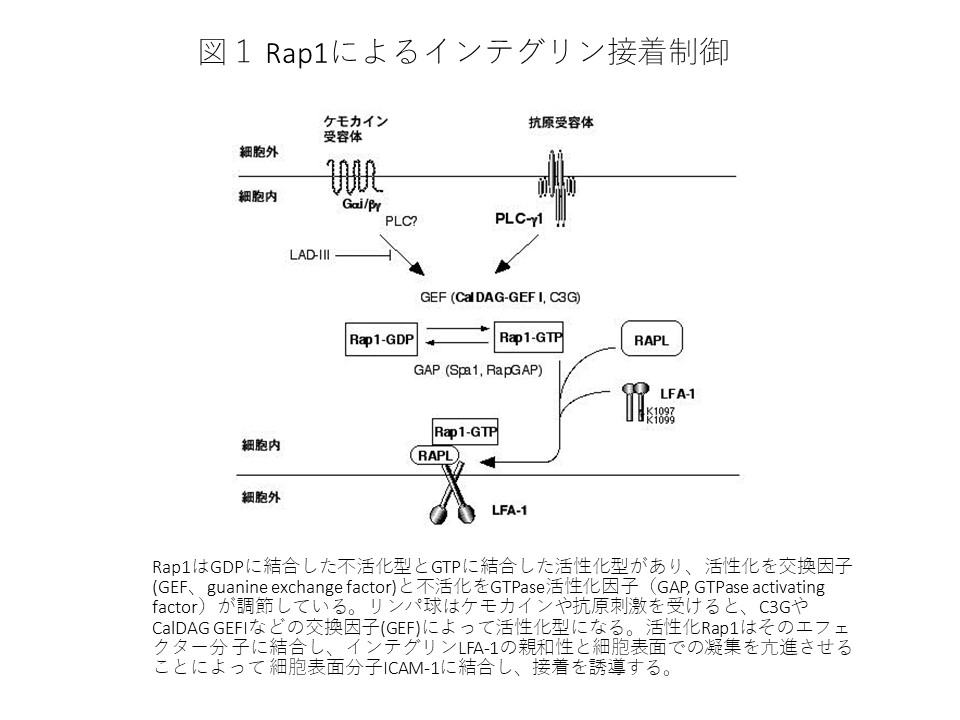
リンパ球は未刺激では浮遊している細胞で血管内を流動していますが、特定のリンパ節や炎症部位に至るとケモカイン刺激によって速やかに血管内皮に接着し、組織へ遊走していきます。この過程でインテグリンであるLFA-1やVLA-4はケモカイン刺激によって発現を変化させず急速に接着性が亢進し、その結果血管内皮に発現しているICAM-1やVCAM-1に結合してリンパ球の血管内皮への接着を誘導することが出来ます。またTリンパ球が抗原を認識し免疫応答を引き起こすのに必要な樹状細胞との接着にLFA-1/ICAM-1を介した免疫シナプスと呼ばれる特殊な接着構造が抗原受容体からのシグナルで形成されます。これらの過程のシグナル伝達とそれによって引き起こされるインテグリン接着制御の様式は大きな謎でした。当研究室では低分子量G蛋白質Rap1がこれらの過程を調節する鍵となる分子であることを突き止めました(図1)
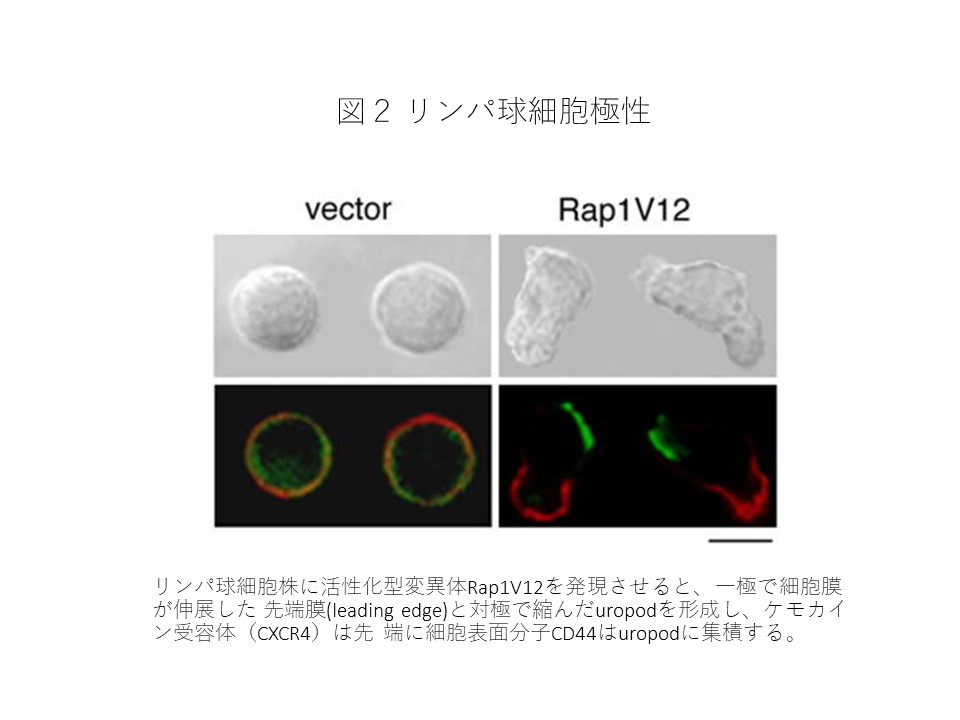
Rap1はインテグリンを介する接着を亢進させるだけでなくリンパ球に前後の細胞極性を与えて活発な細胞移動を引き起こすことが明らかになりました(図2)。
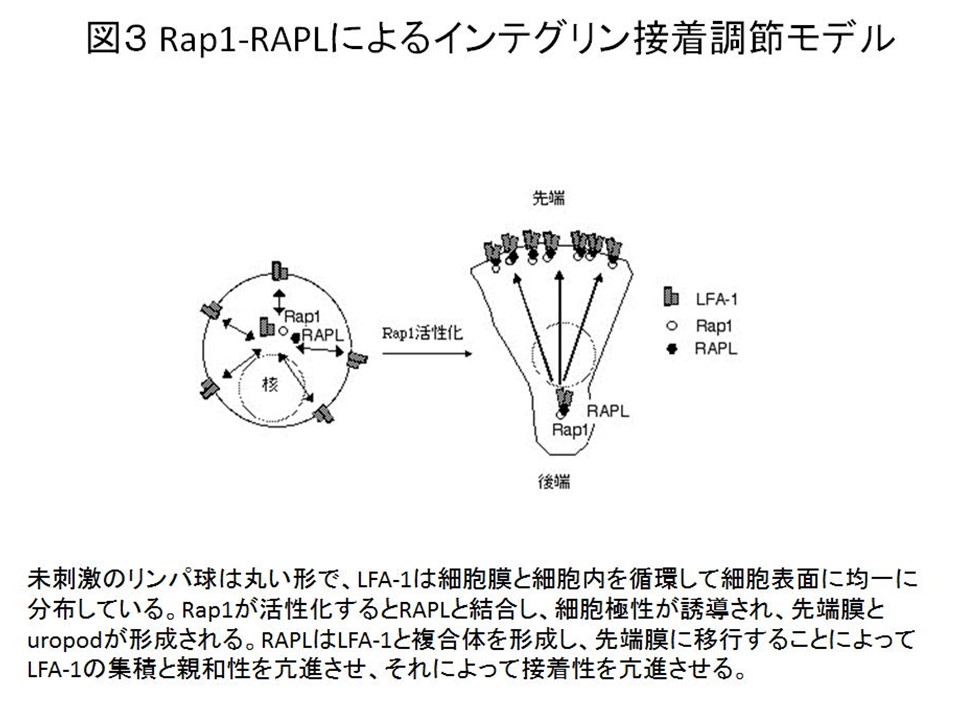
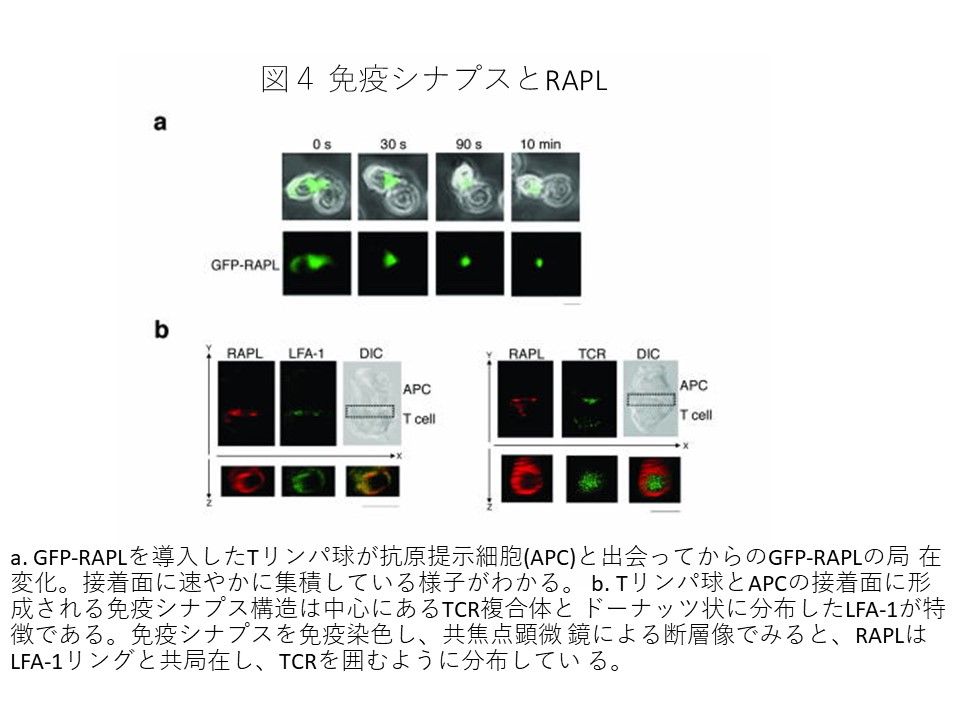
還流下における血管内皮通過過程を顕微鏡下で再現して調べてみると、Rap1が活性化するとリンパ球が血管内皮に接着し、移動通過していくことがわかりました。最近ヒト遺伝病である白血球接着不全症でRap1の活性化不全を伴うケースが見いだしました。このようにRap1が細胞接着、移動に重要な働きをしていることから、Rap1の活性化調節は生体防御において重要と考えられます。 しかしRap1がインテグリンの接着を制御する機構や、その重要性を個体レベルで調べるにはRap1のエフェクター分子を同定しなくてはなりません。当研究室ではインテグリン接着制御を調節する新規Rap1エフェクター分子RAPLを単離しました。解析の結果、RAPLはRap1に結合すると細胞極性を誘導でき、さらにLFA-1と複合体を形成して先端側に集積させる働きをしていることが明らかになりました。(図1,3)。RAPLは抗原呈示細胞と接着すると速やかに接着面に集積し、免疫シナプスを形成する過程に関与します(図4)。
免疫システムは、免疫細胞の動態制御を基盤として成り立っています。顕微鏡技術の発達で生きたマウスのリンパ節内でのリンパ球や樹状細胞の活発な動きや相互作用が観察できるようになりました。このような技術革新は同時に免疫学にその動きや接着の意味を問いかけています。すなわち生体内でリンパ球や樹状細胞の生体内動態変化や細胞間接着を介したコミュニケーションが如何に制御され、その制御が免疫システムのホメオスターシスや適切な免疫応答にどのような役割をはたしているのかを明らかにすることは、現在免疫学が突きつけられている大きな課題といっても過言ではないでしょう。当研究室ではRap1/RAPLによるインテグリン接着制御、細胞極性や細胞遊走の分子機構の解明を行い、これを突破口にして、これらの重要課題に取り組んでいきます。Rap1/RAPLシグナルによるリンパ球や樹状細胞などの免疫細胞の動態と抗原応答の仕組みを個体レベルで調べ、免疫系への影響を探索します。これらの研究を通じて、免疫系の破綻である自己免疫病のあらたな姿が見えてくると予想しています。さらにRAPL欠損マウスの解析から細胞接着と細胞増殖との予期しなかった関係も見えてきました。これはRap1/RAPLシグナルによる接着機能が増殖制御という大きな枠組みで調整されていることを示唆しています。接着研究のパラダイムを変えることになるかもしれないと“わくわく、どきどき”しながら研究を進めています。