丂
PI3K宱楬偲偼
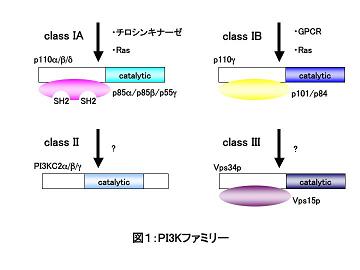
PI3K乮phosphatidylinositol-3 kinase乯偼丄枌偺峔惉惉暘偱偁傞僀僲僔僩乕儖儕儞帀幙偺僀僲僔僩乕儖娐俁埵偺儕儞巁壔傪攠夘偡傞帀幙僉僫乕僛偱偁傝丄傎擕椶偱偼僋儔僗IA丄僋儔僗IB丄僋儔僗II丄僋儔僗III偺係偮偺僒僽僋儔僗偵暘椶偝傟傞乮恾侾嶲徠乯丅
僋儔僗IA PI3K偼丄SH2傪桳偡傞惂屼僒僽儐僯僢僩乮p85α丄p55α丄p50α丄p85β丄p55γ乯偲怗攠僒僽儐僯僢僩乮p110α丄p110β丄p110γ乯偐傜側傞擇検懱偲偟偰懚嵼偟丄TCR傗BCR丄FcεRI偺僔僌僫儖偵墳摎偟偰丄僠儘僔儞僉僫乕僛壓棳偺僔僌僫儖揱払宱楬傗Ras偵傛偭偰妶惈壔偝傟傞丅
惂屼僒僽儐僯僢僩偺撪丄p85α丒p55α丒p50α偼摨堦偺堚揱巕乮Pik3r1乯偺splice variant偱偁傝丄柶塽宯偺嵶朎偱偼庡偲偟偰p85α偑敪尰偟偰偄傞丅
柺敀偄偙偲偵丄PI3K偺惂屼僒僽儐僯僢僩偼怗攠僒僽儐僯僢僩偺僞儞僷僋幙儗儀儖偱偺埨掕壔傪傕偨傜偡乬僔儍儁儘儞乭偲偟偰偺婡擻傕帩偭偰偍傝丄惂屼僒僽儐僯僢僩偺堚揱巕攋夡偵傛偭偰怗攠僒僽儐僯僢僩偺敪尰儗儀儖偺掅壓偑堷偒婲偙偝傟傞丅
怗攠僒僽儐僯僢僩偼嵶朎偺庬椶偵傛偭偰巊偄暘偗偑側偝傟偰偍傝丄奺庬偺堚揱巕夵曄儅僂僗偺夝愅偐傜丄儕儞僷媴丒儅僗僩嵶朎偵偍偄偰偼摿偵p110δ偑庡梫側婡擻傪扴偭偰偄傞傕偺偲峫偊傜傟偰偄傞丅
僋儔僗IB PI3K傕怗攠僒僽儐僯僢僩p110γ偲惂屼僒僽儐僯僢僩乮p101丄p84乯偐傜側傞擇検懱偲偟偰懚嵼偡傞偑丄GPCR乮G-protein coupled receptor乯偺壓棳偺Gβγ傕偟偔偼Ras偵傛偭偰妶惈壔偝傟傞丅
僋儔僗IB PI3K偼敀寣媴偵摿堎揑偵敪尰偟偰偍傝丄働儌僇僀儞偵墳摎偟偨嵶朎梀憱偵娭梌偡傞丅
僋儔僗II丒僋儔僗III PI3K偺柶塽宯偵偍偗傞婡擻偵娭偟偰偼枹偩晄柧側揰偑懡偄偑丄僋儔僗III PI3K偑庬傪挻偊偰嵶朎撪彫朎桝憲偵娭梌偡傞偙偲偐傜丄庬乆偺婷怘夁掱傊偺娭梌偑帵嵈偝傟傞丅
慡偰偺僋儔僗偺PI3K妶惈傪慾奞偡傞LY294002傗Wortmannin偵壛偊丄IC87114乮p110δ摿堎揑乯傗AS605240乮p110γ摿堎揑乯側偳偺奺怗攠僒僽儐僯僢僩偵崅偄慖戰惈傪桳偡傞慾奞嵻偺奐敪偑恑傫偱偄傞丅
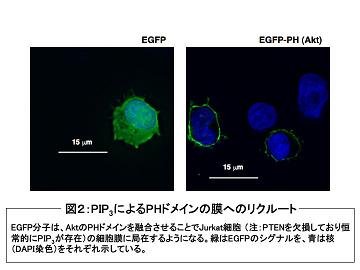
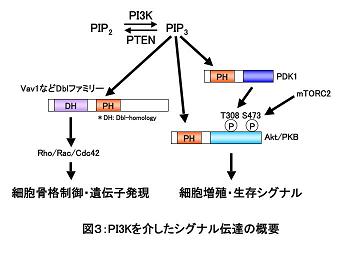
妶惈壔偟偨僋儔僗IA丒IB PI3K偼丄PI(3,4,5)P 俁 (phosphatidylinositol(3,4,5)-trisphosphate) 傗PI(3,4)P俀 (phosphatidylinositol(3,4)-bisphophate)偺惗惉傪夘偟偰丄PH僪儊僀儞乮pleckstrin homology domain乯傪桳偡傞暘巕傪枌傊偲儕僋儖乕僩偡傞乮恾俀嶲徠乯丅
椺偊偽丄枌偵儕僋儖乕僩偝傟偨僙儕儞乛僗儗僆僯儞僉僫乕僛Akt乮PKB偲傕屇偽傟傞乯偼丄摨偠偔PH僪儊僀儞傪桳偡傞PDK1偲丄mTOR丒rictor丒mLST8丒mSIN1偐傜峔惉偝傟傞mTORC2暋崌懱偵傛傝丄偦傟偧傟Thr308偲Ser473偺儕儞巁壔傪庴偗偰妶惈壔偟丄GSK3傗BAD丄FOXO丄MDM2摍偺儕儞巁壔傪夘偟偰嵶朎憹怋傗惗懚僔僌僫儖傪惂屼偡傞乮恾俁嶲徠乯丅
PI3K偼丄懠偵傕丄Vav傪戙昞偲偡傞Dbl僼傽儈儕乕暘巕傪夘偟偰嵶朎崪奿惂屼偵娭梌偡傞偙偲偑抦傜傟傞丅
偑傫梷惂堚揱巕嶻暔偱偁傞PTEN乮phosphatase and tensin homolog deleted on chromosome 10乯偼丄僀僲僔僩乕儖娐偺俁埵偺儕儞巁婎傪扙儕儞巁壔偡傞帀幙儂僗僼傽僞乕僛偲偟偰PI3K宱楬偵漢峈偡傞乮恾俁嶲徠乯丅
幚嵺丄PTEN偺寚幐偼PI(3,4,5)P俁傗PI(3,4)P俀偺憹壛傪夘偟偰峆忢揑側Akt偺妶惈壔傪傕偨傜偟丄Notch傗Ras-MAPK宱楬偑PTEN偺揮幨梷惂傪夘偟偰娫愙揑偵PI3K宱楬偺妶惈壔偺槾恑偵婑梌偟偰偄傞壜擻惈傕帵嵈偝傟偰偄傞丅
偛偔嵟嬤丄PTEN暘巕偺埨掕惈傗嬊嵼偑丄儕儞巁壔傗儐價僉僠儞壔傪夘偟偰惂屼偝傟偰偄傞偙偲偑帵偝傟偨丅
丂
丂